Spinnaker Cove, Long Beach, California
The moon jellyfish Aurelia sp.1 has the largest geographic range
of all species of Aurelia studied to date. It occurs, at least,
in Tokyo Bay and northern Japan, in Australia, on the Atlantic and Mediterranean
coasts of France, and from Los Angeles to San Diego (Dawson & Jacobs
2001; Schroth et al. 2002; Dawson 2003); it probably also exists in
San Francisco Bay (Greenberg et al. 1996; Dawson & Martin 2001).
Each of the other 10+ known species of moon jellyfish have more limited
geographic ranges. For example, A. aurita occurs in the cold-temperate
North Atlantic, A. labiata in the cold-temperate eastern North
Pacific, and A. limbata in the northern polar oceans. Populations
of Aurelia sp. 1 thus appear to be disjunct, while populations
of other species of moon jellyfish generally are contiguous (below).
Comparison of genetic variation, however, shows a very different pattern.
Genetic variation in Aurelia sp.1 is smaller than genetic variation
in most other species of Aurelia (Schroth et al. 2002; Dawson
2003).
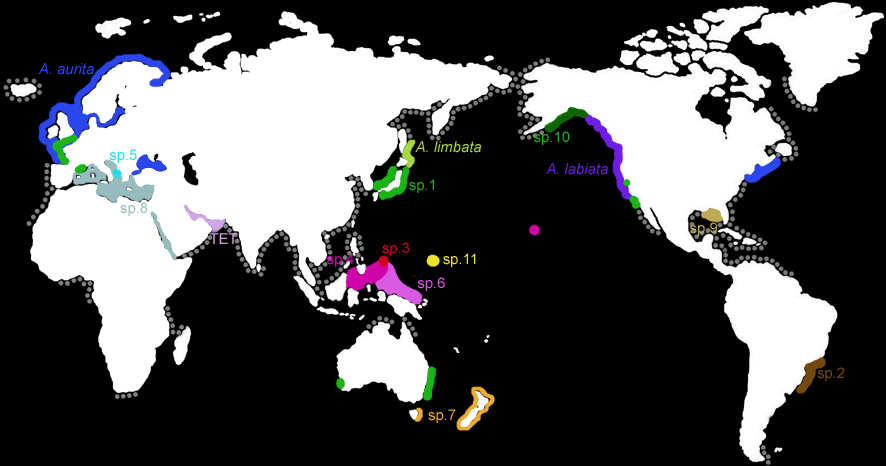
The global distribution
of the genus Aurelia (grey) and the known ranges of species
(colors and
letters, as indicated by molecular and morphological analyses).
The ranges of most species are still poorly defined.
While there are several possible explanations for the observed contrast, such low genetic variation coupled with such a large geographic range that includes disjunct populations in some of the major warm-temperate seaports of the world suggests that Aurelia sp. 1 may be an introduced species. Notably, the latitudinal, or temperature, range over which Aurelia sp. 1 occurs is similar in each of the five major coastal regions from which it is currently known, a pattern that is commonplace in exotic species and may reflect underlying ecological processes (Sax 2001). Although its point of origin and the pattern and timing of introductions are currently unknown, wider geographic sampling and more extensive molecular analyses could clarify these issues (e.g. Holland 2000).

Aurelia
sp.1
Japan
Species introductions pose a major threat to biodiversity (Lee 2002). Jellyfish, like other coastal marine species, can be accidentally introduced as organisms growing on ship hulls or in ballast water, or released unintentionally (or intentionally) as a result of the aquarium trade and aquaculture (Holland 2000; Grosholz 2002). The scale of species introductions is bewildering. For example, the USA receives between 20 million and 80 million tonnes of ballast water per year that is laced with an average of one billion bacteria per litre from foreign ports (Ruiz et al. 2000; ANS). The numbers are smaller for larger organisms, and only a subset become established, but the numbers are still impressive. At least 287 introduced, or 'exotic', marine species have become established in Hawai'i (Eldredge & Carlton 2002), between 212-335 in San Francisco Bay (USGS), and 120 in the Chesapeake Bay (ANS). Moreover, their effect is disproportionately large. Over 15% of exotic species cause serious harm, exotic species negatively impact at least 42% of endangered species, and the cost associated with major exotic species in the USA alone is on the order of US$100 billion per year (Lee 2002; ANS).

Aurelia sp.1
Australia
Causative
links between introduced species of jellyfish (scyphozoans, hydrozoans,
ctenophores), increases in the occurrence or severity of jellyfish blooms,
and decreases in the abundance of endemic gelatinous zooplankton can
still be somewhat obscure (Mills 2001). This may be due in part to the
existence of 'cryptogenic' species, i.e. species whose endemic or exotic
status is unclear (Carlton 1996; Mills 2001) and in part to difficulties
in ascribing causal relationships when other influences such as eutrophication
are also important (Mills 2001). However, several candidates are likely
to be nuisance exotic scyphozoans. For example, Rhopilema nomadica
was first recorded in the eastern Mediterranean in 1976, and has since
become abundant off Israel each summer; it is hazardous to bathers and
fishermen due to its sting and may also clog fishing nets. Presumably
exotic Aurelia sp. have clogged seawater intakes and shut-down
power plants in Australia, the Baltic region, India, Korea, Philippines,
and Saudi Arabia (Mills 2001). Such blooms may be facilitated by small
ecological advantages conferred on the exotics by 'genotype x environment'
interactions (Dawson & Martin 2001). Other exotic scyphomedusae
that have had negative impacts in recent years include Phyllorhiza
punctata, in the Mediterranean and Gulf of Mexico, and Cassiopea
spp., in the Mediterranean and Hawai'i (Devaney & Eldredge 1977;
Mills 2001; Graham et al. 2003; Holland et al. unpubl.).